
Home / Journal / Endocrinology, Diabetes and Obesity
Special Issue: Tissue- and Cell-Specific Dysregulations of ACE2 may put Patients with Diabetes at Higher Risk for COVID-19
SARS-Coronavirus-2 Diabetes Metabolic Syndrome Inflammation
Marko Marhl
DOI: 10.31532/EndocrinolDiabetesObes.2.1.001 03 Nov 2020
Download

Abstract
Hypertension and diabetes are the major risk factors for patients with Coronavirus disease 2019 (COVID-19). The physiological link between the diseases is still obscure. A growing body of evidence indicates that Angiotensin-converting enzyme 2 (ACE2) could be one of the critical agents linking diabetes with hypertension and COVID-19. Whether an up- or rather a down-regulation of ACE2 is responsible for the higher risk for COVID-19 is not clear. Experimental evidence and theoretical models exist for both. We show here that in addition to the overall up-/down-regulation of ACE2 at the whole-body level, we need to consider the tissue- and cell-specific dysregulations of ACE2, determining the risk of patients with diabetes for COVID-19.
Keywords
SARS-Coronavirus-2, Diabetes, Metabolic Syndrome, Inflammation.
Abbreviations
COVID-19: Coronavirus disease 2019; SARS-CoV-2: Severe Acute Respiratory Syndrome - Coronavirus-2; ACE2: Angiotensin-converting enzyme 2; sACE2: soluble ACE2; ACEi: Angiotensin-Converting Enzyme inhibitors; ARBs: Angiotensin Receptor Blockers; T2DM - Type 2 Diabetes Mellitus; CVD - Cardio vascular disease; NAFLD - Non-alcoholic fatty liver disease; DCM: Dilated cardiomyopathy; HCM: Hypertrophic cardiomyopathy; TMPRSS2: Transmembrane protease serine 2.
Highlights
• Hypertension and diabetes are the major risk factors for patients with COVID-19.
• Dysregulation of ACE2 expression links hypertension and diabetes with COVID-19.
• Tissue- and cell-specific ACE2 dysregulations better determine the risk for COVID-19 then an overall whole-body up- or down-regulation of ACE2 expression.
Introduction
The epidemic of Coronavirus disease 2019 (COVID-19), caused by the Severe Acute Respiratory Syndrome - Coronavirus-2 (SARS-CoV-2), is rapidly spreading and affecting millions of people all over the world. Clinical studies have shown that hypertension and diabetes represent the leading risk factors for COVID-19 1. The underlying physiological processes linking these comorbidities with COVID-19 are still obscure. The relationship between diabetes and COVID-19 is incredibly complex, and several studies have contributed to a better understanding of how diabetes is linked to more severe COVID-19 illness and death, for review 2-4. As diabetes, in particular Type 2 Diabetes Mellitus (T2DM), is strongly associated with elevated adipose tissue mass, obesity with a high BMI might be an essential risk factor for a severe course of COVID-195. However, not merely the BMI reflects the risk, but the body fat distribution may be a better proxy for obesity-related health risks. Novel findings suggest an independent role of increased gluteofemoral fat mass to maintain metabolic health, and the risk of visceral adiposity might be equally important as the risk of a lower amount of lower-body fat mass6. Interestingly, once the patients with COVID-19 are in the intensive care unit, obesity is known to confer a survival advantage, the so-called “obesity paradox”. Patients with BMI>25 kg⁄m2 have a better chance to survive mechanical ventilation and severe septic states than patients with a normal or low BMI.7
The relationship between T2DM and obesity with COVID-19 is multifactorial and may also be bi-directional. One of the main risks of obesity and adipocyte dysfunction is a low persistent inflammation, i.e., meta-inflammation.7 Adipocytes and immune cells act cooperatively to produce cytokines and chemokines that mobilize the rapid recruitment of inflammatory macrophages.8 The immune cells interaction in adipose tissue in obesity is related to the macrophage shift, and the circulating inflammatory markers, including cytokines and chemokines, are often related to non-alcoholic fatty liver disease (NAFLD). 9 An excessively activated immune response may contribute to a severe COVID-19. The host inflammatory response can reach the hyperinflammatory phase, as viral levels already decline.10 The hyperreactivity of the immune system can result in adipocyte inflammation, NAFLD, and T2DM.11 The bi-directional relationship between diabetes and COVID-19 has been discussed 12, and was also reported for SARS CoV, showing that SARS CoV can enter the pancreas and damage islets, causing acute diabetes.13
In our previous work, data mining analysis of publications in PubMed has revealed three main routes linking diabetes and COVID-19 via systemic inflammation, liver dysfunction, and dysregulated Angiotensin-Converting Enzyme 2 (ACE2) expression.14 The dysregulation of ACE2 has attracted interest by several researcher groups because ACE2 serves as the cellular entry point for virus SARS-CoV-2.3,15,16 In an early phase of T2DM, ACE2 might be upregulated,17, 18 and could be an adaptive response compensating the low-grade systemic inflammation characterizing T2DM,19 as ACE2 is known as an effective anti-inflammatory and antifibrogenic agent20. In a later phase of T2DM, however, the ACE2 might be downregulated,18 which associates with several complications, e.g., diabetic nephropathy, oxidative stress in the pancreas, and impaired insulin secretion. 17,21
Here we discourse the influence of ACE2 dysregulations on the course of COVID-19. We link the ACE2 dysregulations to age and chronic diseases that might potentiate the risk for a more severe course of COVID-19. In particular, T2DM is in the main focus of this study. We compare the importance of an overall whole-body ACE2 dysregulation with the tissue- and cell-specific ACE2 overexpression.
Whole-body ACE2 expression
The consequences of a whole-body downregulation of ACE2 expression in patients with a developed T2DM are not entirely understood.18 On the one hand, ACE2 plays a protective role because of its anti-inflammatory role,7,20 however, on the other hand, it represents the virus entry point, and a dilemma arises what the net effect of this dysregulation in ACE2 expression is. For elderly patients, it is even more challenging because the whole-body level ACE2 expression might decrease with age,22 and this downregulation appears to be accelerated in patients with T2DM.23,24 Therefore, it might be speculated that a whole-body downregulation of ACE2 expression could explain the higher mortality for COVID-19 in elderly patients with T2DM.
The link between the risk for severity of COVID-19 and the whole-body ACE2 expression is not so trivial. It should be noted that other researchers couldn’t find any statistically relevant differences in the whole-body ACE2 expression related to age.25 The diverse outcomes could be explained by noting that the expression of ACE2 differs in different organs and tissues. The ACE2 expression can be upregulated in one part of the body and downregulated in the others without any net-effect on the whole-body ACE2 expression. Indeed, Xu et al.26 have shown that the expression of ACE2 in the lung increases with age, and they also showed that an up-regulation in a particular tissue depends on the grade of injury. In gastric tissue, for example, ACE2 expression was gradually increased from chronic gastritis, metaplasia to early cancer26 . In the lung, elevated expression of ACE2 was found in cigarette smokers. 27,28 It has been speculated that long-term smoking may increase the risk for COVID-19 because ACE2 is not only a receptor but is also involved in post-infection regulation, including immune response, cytokine secretion, and viral genome replication.28
The systemic whole-body regulation of ACE2 expression might be misleading when related to pathologies, and in particular, in COVOD-19, it can lead to diverse, often contradictory, conclusions. This interrelation between the diseases is additionally challenging when patients with diabetes and/or hypertension are treated with Angiotensin-Converting Enzyme inhibitors (ACEi) and Angiotensin Receptor Blockers (ARBs), the known drugs for influencing the ACE2 expression.3,4,19,29 Therefore, we focus on the tissue- and cell-specific ACE2 expression that can give better insight into the pathology of COVID-19 and its relationship with the most common comorbidities, i.e., T2DM, CVD, and obesity.
Tissue- and cell-specific upregulation of ACE2 expression
ACE2 is differently expressed among tissues,30 and the distribution of ACE2 changes with age and in dependence on the stage of disease.26 In particular, chronic diseases might shift the ACE2 expression considerably.3 T2DM, characterized by chronic low-grade inflammation, influences the distribution of ACE2, and this might not only be at the whole-body level,18 but it is usually tissue specific. Wysocki et al.31 have shown experimentally for db/db and db/m models of diabetic mice that the ACE2 activity is upregulated explicitly in the renal cortex. Renal failure is a severe complication of COVID-19. Due to the known anti-inflammatory role of ACE2, 7,20 it might be speculated that ACE2 is upregulated in the tissues mostly affected by the low-grade inflammation caused by the comorbid chronic disease. Therefore, the tissue-dependent upregulations in ACE2 expression indicate a link between diabetes, chronic inflammation, and ACE2 in patients with COVID-19.
Notably, the expression of ACE2 might not only be tissue-dependent, but it may considerably differ among the cells in a given tissue. For example, in diet-induced obese mice, but not in lean mice, a higher expression of ACE2 was explicitly observed in the lung epithelial cells. In the human lungs, ACE2 was also overexpressed exclusively in the epithelial cells. 32 This might be of particular importance in the epidemic of COVID-19 because several fatal cases are linked to damages in the lung epithelial tissue. Kruglikov and Scherer33 argue that ACE2 is overexpressed in adipocytes and adipocyte-like cells, such as pulmonary lipofibroblasts. The expression of ACE2 is upregulated in adipocytes of patients with obesity and diabetes. The pulmonary lipofibroblasts potentially transdifferentiate into myofibroblasts that can lead to pulmonary fibrosis and significantly increase the severity of COVID-19. In verifying this hypothesis, they show that thiazolidinediones, the well-known anti-diabetic drugs, can stabilize lipofibroblasts, preventing the transition to myofibroblasts, reducing the development of pulmonary fibrosis, and thereby lowering the risk for severity of COVID-19. 33
Assessing ACE2 expression in normal and diseased human myocardial tissues profiled by bulk and single nucleus RNA-seq, the bulk RNA-seq data from these individuals show no significant alterations in ACE2 expression in the context of dilated or hypertrophic cardiomyopathy (DCM and HCM, respectively) compared to non-failing controls.34 However, single nucleus RNA-seq highlighted a stark upregulation in ACE2 expression in cardiomyocytes in DCM and HCM, and a concomitant downregulation of ACE2 expression in fibroblasts, pericytes, and vascular smooth muscle. Along the line of this study, Guo et al.35 have also found that ACE2 expression in cardiomyocyte of heart failure samples was higher than in the normal heart. These results suggest that cardiovascular disorders are predominant drivers of cardiomyocyte-specific increased transcription of ACE2.
In another study, high expression of ACE2 was revealed in pericytes of patients with basic heart failure disease,30 The increased ACE2 expression was found at both mRNA and protein levels. Furthermore, He et al.36 have found that ACE2 is specifically and highly expressed in microvascular pericytes of the heart and brain, but not in the endothelial cells of the same tissue. Interestingly, mice with pericyte ablation showed increased expression and release of Von Willebrand Factor from microvascular endothelial cells, which indicates that pericytes regulate thrombogenic responses in neighboring endothelial cells. This finding, identifying pericytes rather than endothelial cells as the cells with high ACE2 expression in the microvasculature, may explain why diabetes, hypertension, and obesity are risk factors for severe COVID-19 patients, as these comorbidities are characterized by an impaired endothelial barrier function, allowing SARS-CoV-2 to reach and infect the pericytes that are normally shielded from the blood behind an intact endothelial barrier.36 The effect of diabetes on endothelial barrier function is well established, and there is evidence that endothelial permeability is increased in obesity.37 The tissue- and cell-specific ACE2 overexpression characterizing particular pathologies, recognized as COVID-19 comorbidities, are schematically presented in Figure 1.
Figure 1. Tissue- and cell-specific ACE2 overexpression associated with COVID-19 comorbidities.
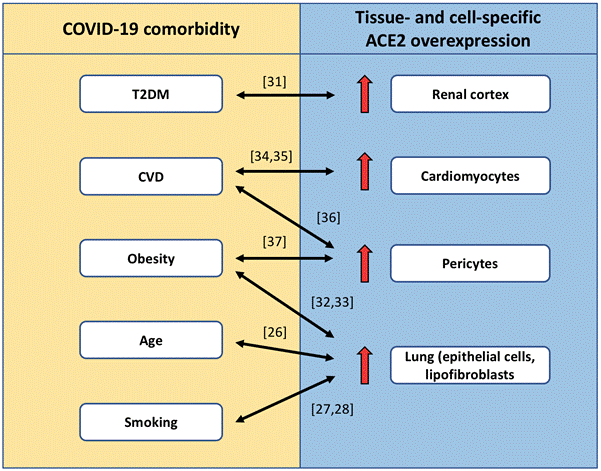
A dysregulated expression of ACE2 among cells and tissues might explain the reasons for the vulnerability of patients with diabetes and other chronic diseases for COVID-19. The cells with an upregulated expression of ACE2 enhance the probability for entry of SARS-CoV-2 and hence increase the risk of infection. Figure 1 shows that the leading COVID-19 comorbidities (T2DM, obesity, CVD including hypertension, age, and smoking) specifically upregulate ACE2 expressions in the most often attacked tissues and cells in COVID-19 (lung and particularly the lung epithelial cells, cardiomyocytes, vasculature and particularly the pericytes, and the renal cortex). In Fig. 1, the references are added to the known relationships between COVID-19 comorbidities and the corresponding tissue- and cell-specific ACE2 overexpression. However, it should be noted that the interconnections presented in Fig. 1 are probably even more complex and will need to be amended in the course of further investigations.
Here we are dealing with the risk of a tissue- and cell-specific ACE2 up-regulation. However, the risk of tissues and cells with a down-regulated ACE2 expression should also be considered. Concerning the pathology of COVID-19, the cells with less ACE2 might be associated with a weaker anti-infection ability of the tissue, and the activation of the inflammatory response induced by SARS-CoV-2 infection.35 Therefore, a fine-tuned regulation of ACE2 expression among the cells and tissues is vital for effective prevention and control of COVID-19.
Conclusions
A dysregulated ACE2 expression in the body might represent a higher risk for COVID-19. Several age-related pathologies are related to ACE2 dysregulations. We provide evidence that the tissue- and cell-specific overexpression of ACE2 enables a better understanding of the relationship between COVID-19 and its comorbidities than considering changes in the whole-body ACE2 expression. The data collected in Fig. 1 shows that T2DM, together with obesity, CVD including hypertension, age, and smoking, specifically upregulate ACE2 expression in tissues and cells of those organs that are most frequently a target of injury in COVID-19 (lung, heart, vasculature, and kidneys). The reality is that patients with T2DM are often obese and hypertense. Therefore, in particular, elderly and obese patients with T2DM are at elevated risk of dysregulated ACE2 expression and a more severe form of COVID-19. Smoking will potentiate this risk.
In the process of aging and developing age-related pathologies, the cell-specific ACE2 expression in different tissues might follow a redistribution of ACE2 to places of a higher tissue injury caused by an underlying chronic disease. In particular, T2DM, systemic disease with an underlying low-grade chronic inflammation, might contribute considerably to the pathological dysregulations of ACE2 expression among the cells in different tissues. Also, other chronic diseases with inflammatory and fibrotic scarring effects might cause tissue injuries with a consequent pathophysiological dysregulation of ACE2 expression, typically with an up-regulation of ACE2 in one part of the cells and a concomitant down-regulation in other cell types of the tissue.34, 35
It should be noted, however, that several other factors might also influence the role of ACE2 in explaining the risk factors for COVID-19. Regarding the ACE2 as a receptor for SARS-CoV-2, it has been found that enzyme Transmembrane protease serine 2 (TMPRSS2) was only expressed in a subset of ACE2+ cells,38 which suggests that SARS-CoV-2 might use alternative pathways for entering the cells. It has indeed been shown that SARS-CoV-2 could also enter TMPRSS2− cells using cathepsin B/L. 39
In this paper, we have focused on the membrane-bound ACE2, but it should be pointed out that the soluble ACE2 (sACE2) might also play an essential role in the pathology of COVID-19.40 Because sACE2 can interact directly with SARS-CoV-2, sACE2 could potentially act as a virus trap and its effective inactivator40–42 . Therefore, a new medical treatment was proposed by implementing a recombinant sACE2 that would inhibit the infection with SARS-CoV-2 by shifting the competition with membrane-bound ACE2 toward the sACE2 that cannot promote viral entry into the cell. 40–42 Although it sounds promising, further studies will be needed to understand better the comprehensive role of ACE2 in COVID-19 and its comorbidities.
Acknowledgements
This work was supported by the Slovenian Research Agency (Research Core Funding, No. P1-0055).
References
- Wang B, Li R, Lu Z, Huang Y. Does comorbidity increase the risk of patients with COVID-19: Evidence from meta-analysis. Aging. 2020; 12, 6049–6057, doi: 10.18632/aging.103000.
- Hussain A, Bhowmik B, do Vale Moreira NC. (2020) COVID-19 and diabetes: Knowledge in progress. Diab Res Clin Pract. 2020;162, 108142, https://doi.org/10.1016/j.diabres.2020.108142.
- Gupta A, Madhavan MV, Sehgal K, et al. Extrapulmonary manifestations of COVID-19. Nat Med. 2020;26, 1017–1032, https://doi.org/10.1038/s41591-020-0968-3 .
- Apicella M, Campopiano MC, Mantuano M, et al. COVID-19 in people with diabetes: understanding the reasons for worth outcomes. Lancet Diabetes Endocrinol. 2020;8, 782–792, https://doi.org/10.1016/ S2213-8587(20)30238-2.
- Stefan N, Birkenfeld AL, Schultze MB, Ludwig DS. Obesity and impaired metabolic health in patients with COVID-19. Nat Rev. 2020;16, 341–342, https://doi.org/10.1038/ s41574-020-0364-6.
- Stefan N. Causes, consequences, and treatment of metabolically unhealthy fat distribution. Lancet Diabetes Endocrinol. 2020;8, 616–627,https://doi.org/10.1016/S2213-8587(20)30110-8.
- Méry G, Epaulard O, Borel A-L, Toussaint B, Le Gouellec A. COVID-19: Underlying Adipokine Storm and Angiotensin 1-7 Umbrella. Front Immunol. 2020;11,1714. doi:10.3389/fimmu.2020.01714.
- Lu J, Zhao J, Meng H, Zhang X. Adipose Tissue-Resident Immune Cells in Obesity and Type 2 Diabetes. Front Immunol. 2019; 10, 1173. doi: 10.3389/fimmu.2019.01173.
- Liu R and Nikolajczyk BS. Tissue Immune Cells Fuel Obesity-Associated Inflammation in Adipose Tissue and Beyond. Front Immunol. 2019; 10, 1587. doi: 10.3389/fimmu.2019.01587.
- Ayres JS. A metabolic handbook for the COVID-19 pandemic. Nat Metab. 2020; 2, 572–585, https://doi.org/10.1038/s42255-020-0237-2 .
- Eigentler T, Lomberg D, Machann J, Stefan N. Lipodystrophic Nonalcoholic Fatty Liver Disease Induced by Immune Checkpoint Blockade. Ann Intern Med. 2020;172, 836–837, doi: 10.7326/L19-0635.
- Rubino F, Amiel SA, Zimmet P, et al. New-Onset Diabetes in Covid-19. N Engl J Med. 2020.DOI: 10.1056/NEJMc2018688.
- Yang J-K, Lin S-S, Ji X-J, Guo L-M. Binding of SARS coronavirus to its receptor damages islets and causes acute diabetes. Acta Diabetol. 2010;47, 193–199, DOI 10.1007/s00592-009-0109-4.
- Marhl M, Grubelnik V, Magdič M, Markovič R. Diabetes and metabolic syndrome as risk factors for COVID-19. Diabetes Metab Syndr Clin Res Rev. 2020;14 (4), 671–477, doi: 10.1016/j.dsx.2020.05.013.
- Xu H, Zhong L, Deng J, Peng J, Dan H et al. (2020) High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int J Oral Sci. 2020;12 (8), 1–5, https://doi.org/10.1038/s41368-020-0074-x.
- Muniyappa R, Gubbi S. COVID-19 Pandemic, coronaviruses, and diabetes mellitus. Am J Physiol Endocrinol Metab. 2020;318, E736–E741, https://doi.org/10.1152/ajpendo.00124.2020.
- Batlle D, Soler MJ, Ye M. ACE2 and Diabetes: ACE of ACEs? Diabetes. 2010;59 (12), 2994–2996, doi: 10.2337/db10-1205.
- Patel VB, Parajuli N, Oudit GY. Role of angiotensin-converting enzyme 2 (ACE2) in diabetic cardiovascular complications. Clin Sci (Lond). 2014;126 (7), 471–482, doi: 10.1042/CS20130344.
- Pal R, Bhansali A. COVID-19, diabetes mellitus and ACE2: The conundrum. Diab Res Clin Pract. 2020;162, 108132, doi: 10.1016/j.diabres.2020.108132
- Prestes TRR, Rocha NP, Miranda AS, et al. The anti-inflammatory potential of ACE2/Angiotensin-(1-7)/Mas receptor axis: Evidence from basic and clinical research. Curr Drug Targets. 2017;18 (11), 1301–1313, doi: 10.2174/1389450117666160727142401.
- Graus-Nunes F, Souza-Mello V. The renin-angiotensin system as a target to solve the riddle of endocrine pancreas homeostasis. Biomed Pharmacother. 2019;109, 639–645, doi: 10.1016/j.biopha.2018.10.191.
- Yoon HE, Kim EN, Kim MY, et al. (2016) Age-Associated Changes in the Vascular Renin-Angiotensin System in Mice. Oxid Med Cell Longev. 2016, 6731093, http://dx.doi.org/10.1155/2016/6731093.
- Ciaglia E, Vecchione C, Puca AA. COVID-19 Infection and circulating ACE2 levels: Protective role in women and children. Front Pediatr. 2020; 8, 206, doi: 10.3389/fped.2020.00206.
- Chen J, Jiang Q, Xia X. et al. Individual variation of the SARS-CoV-2 receptor ACE2 gene expression and regulation. Aging Cell. 2020;19, e13168, https://doi.org/10.1111/acel.13168.
- Li M-Y, Li L, Zhang Y, Wang X-S. Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues. Infect Dis Poverty. 2020;9, 45, https://doi.org/10.1186/s40249-020-00662-x.
- Xu J, Chu M, Zhong F, et al. Digestive symptoms of COVID-19 and expression of ACE2 in digestive tract organs. Cell Death Discovery. 2020;6, 76, https://doi.org/10.1038/s41420-020-00307-w.
- Leung JM, Yang CX, Tam A, et al. ACE-2 Expression in the Small Airway Epithelia of Smokers and COPD Patients: Implications for COVID-19. Eur Respir J. 2020, in press, https://doi.org/10.1183/13993003.00688-2020.
- Li G, He X, Zhang L, et al. Assessing ACE2 expression patterns in lung tissues in the pathogenesis of COVID-19, J Autoimmun. 2020;112, 102463, https://doi.org/10.1016/j.jaut.2020.102463.
- Cavallo F, Chiarelli F. SARS-CoV-2 Infection and Diabetes. Curre Res Diabetes & Obes J. 2020;13(2), 555856, DOI: 10.19080/CRDOJ.2020.13.555856.
- Chen L, Li X, Chen M, Feng Y, Xiong C. The ACE2 expression in human heart indicates new potential mechanism of heart injury among patients infected with SARS-CoV-2. Cardiovasc Res. 2020;116(6, 1097–1100, doi:10.1093/cvr/cvaa078.
- Wysocki J, Ye M, Soler MJ, Gurley SB, Xiao HD et al. ACE and ACE2 Activity in Diabetic Mice. Diabetes. 2006;55, 2132-2139, DOI: 10.2337/db06-0033.
- Al Heialy S, Hachim MY, Senok A, Gaudet M, Tayoun AA et al. Regulation of Angiotensin-Converting Enzyme 2 (ACE2) in Obesity: Implications for COVID-19. Front Physiol. 2020;11, 555039. doi: 10.3389/fphys.2020.555039.
- Kruglikov IL, Scherer PE. The Role of Adipocytes and Adipocyte-Like Cells in the Severity of COVID-19 Infections. Obesity. 2020;28, 1187-1190, doi:10.1002/oby.22856.
- Tucker NR, Chaffin M, Bedi KC, et al. Myocyte specific Upregulation of ACE2 in Cardiovascular Disease: Implications for SARS-CoV-2-Mediated Myocarditis. Circulation. 2020;142, 708–710, https://doi.org/10.1161/CIRCULATIONAHA.120.047911.
- Guo J, Wei X, Li Q, et al. Single-cell RNA analysis on ACE2 expression provides insight into SARS-CoV-2 potential entry into the bloodstream and heart injury, J Cell Physiol. 2020;235, 9884–9894, DOI: 10.1002/jcp.29802.
- He L, Mäe MA, Muhi L, et al. Pericyte-specific vascular expression of SARS-CoV-2 receptor ACE2 – implications for microvascular inflammation and hypercoagulopathy in COVID-19. BioRxiv. 2020. DOI: 10.1101/2020.05.11.088500
- Lockhart SM, O’Rahilly S. When Two Pandemics Meet: Why Is Obesity Associated with Increased COVID-19 Mortality?, Med. 2020;1: 1–10, https://doi.org/10.1016/j.medj.2020.06.005.
- Sungnak W, Huang N, Bécavin C, et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat Med 26 (May 2020): 681–687, https://doi.org/10.1038/s41591-020-0868-6.
- Hoffmann M, Kleine-Weber H, Schroeder S, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020;181, 271–280, https://doi.org/10.1016/j.cell.2020.02.052.
- Batlle D, Wysocki J, Satchell K. Soluble angiotensin-converting enzyme 2: a potential approach for coronavirus infection therapy? Clin Sci. 2020; 134, 543–545, https://doi.org/10.1042/CS20200163.
- Monteil V, Kwon H, Prado P, et al. Inhibition of SARS-CoV-2 infections in engineered human tissues using clinical-grade soluble human ACE2. Cell. 2020;181, 905–913, https://doi.org/10.1016/j.cell.2020.04.004.
- Alhenc-Gelas F, Drueke TB. Blockade of SARS-CoV-2 infection by recombinant soluble ACE2. Kidney Int. 2020;97, 1091–1096, https://doi.org/10.1016/j.kint.2020.04.009